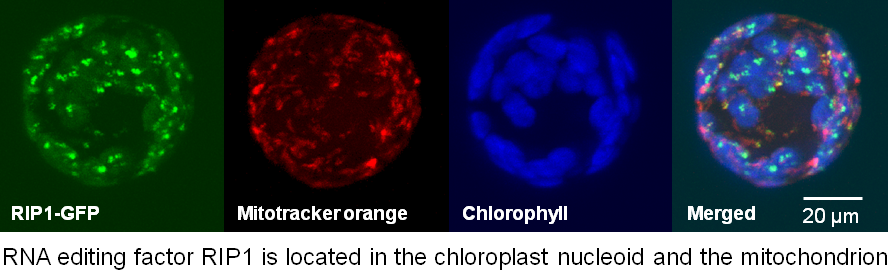
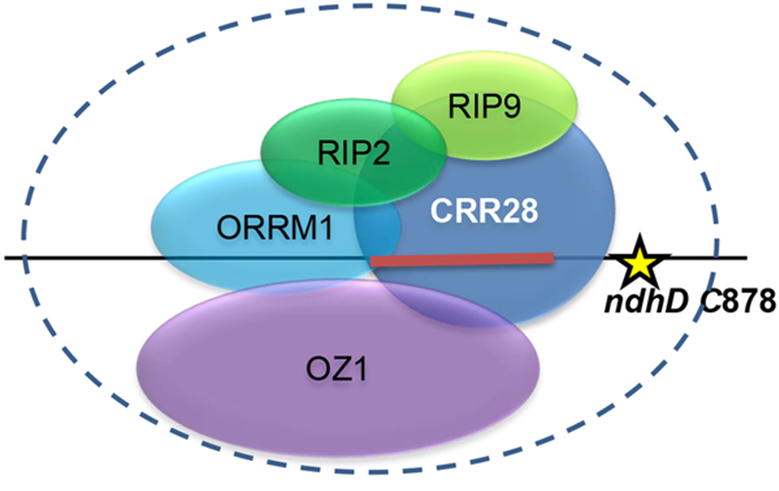
Particular cytidines in transcripts in chloroplasts and mitochondria of plant cells undergo modification to uridine by RNA editing. These C-to-U changes can create start and stop codons or change the encoded amino acid from the one predicted from the genomic sequence. RNA editing is known to occur in a variety of organisms, including mammals, insects, plants and microorganisms. In humans and other animals, RNA editing creates diversity in certain classes of proteins encoded by the nuclear genome. Editing in plants appears to correct defective RNA post-transcriptionally.

We have identified the C targets recognized by several PPR proteins, such as RARE1 and QED1.
Some members of the PPR protein family contain a DYW domain at the C-terminus. This domain has been hypothesized to provide the enzymatic activity needed to convert C to U because it carries motifs characteristic of cytidine deaminases. We performed site-specific mutagenesis of these motifs in two PPR proteins and demonstrated that editing activity was lost. This finding supports the idea that DYW domains carry cytidine deaminase activity.